

Hoe kunnen we helpen?
Viscosin
Introduction
General information
Viscosin is a cyclic lipopeptide produced by various Pseudomonas species. It is the namesake CLiP of the viscosin group, a collection of structurally similar CLiPs. Viscosin can be considered the first discovered Pseudomonas CLiP. It was first described in 1951 for its antiviral properties against human pathogenic viruses such as the infectious bronchitis virus, influenza virus and Newcastle disease virus. (Groupé et al., 1951) Since then, viscosin has been found in a variety of natural environments, including marine and rhizosphere environments.
| Original publication | Groupé et al., 1951 |
| Original source | Pseudomonas viscosae |
| Other known sources (non-putative) | Pseudomonas sp. SBW25, Pseudomonas sp. SH10-3B, Pseudomonas sp. MK91CC8, Pseudomonas libanensis M9-3 |
| Stereochemistry determined by | Chemical synthesis (P. viscosae) Enzymatic deamination (P. sp. SH10-3) Chiral GC (P. sp. MK91CC8) X-ray structure (P. sp. SBW25; unpublished) |
Chemical information
| CAS | 27127-62-4 |
| Molecular formula | C54H95N9O16 |
| Molecular weight | 1126.4 g/mol |
| Mono-isotopic mass | 1125.6897 Da (isobaric to other CLiPs, see text) |
| Solubility | Acetonitrile, chloroform, acetone, dioxane, buffer Not soluble (or very limited) in benzene or miliQwater |
| CMC | 48 µM (Saini, 2008) 3.6 – 8.0 µM (Laycock, 1991) 133 µM (Hildebrand, 1998) |
| Minimal surface tension | 28 mN/m (Saini, 2008) |
| 3D conformation | Solution NMR structure (unpublished) X-ray structure (unpublished) |
| NMR data available in literature | Methanol-d4 (Laycock, 1991) (P. sp. SH10-3B) Acetone-d6 (Gerard et al., 1997) (P. sp. MK91CC8) Methanol-d4 (De Bruijn, 2007) (P. sp. SBW25 |
Biological activity
In general, viscosin displays an antagonistic activity against Gram-positive bacteria, mycobacteria, protozoa and fungi. (Gerard, 1997; De Bruijn, 2007; Geudens, 2017) No information is available on its haemolytic or insecticidal activity. Viscosin also inhibited migration of two metastatic human cancer cell lines (breast and prostate). (Saini, 2008) In the context of activity against Gram-negative bacteria, pure viscosin has only been applied to Escherichia coli, whereby no growth inhibition has been observed. (Gerard, 1997)
Chemical structure
The structure determination of viscosin was not straightforward: Initially, a linear structure composed of 6 amino acid coupled to a fatty acid was proposed. (Ohno, 1953). However, chemical syntheses later revealed that this structure was erroneous. (Hiramoto, 1969) The correct structure of viscosin was only described in 1970, 19 years after its initial discovery. (Hiramoto, 1970) The correct amino acid sequence of viscosin is 3-OH C10:0 – L-Leu1 – D-Glu2 – D-aThr3 – D-Val4 – L-Leu5 – D-Ser6 – L-Leu7 – D-Ser8 – L-Ile9, whereby the molecule is cyclized between the C-terminus and the side chain hydroxyl moiety of threonine at position 3 by the formation of an ester bond. In this way, the molecule is composed of 9 amino acids, 7 of which are involved in the macrocycle. The molecule has a molecular weight of 1126.4 g/mol (molecular formulae C54H95N9O16) which is identical (isobaric) to that of WLIP, massetolide F and massetolide L, all CLiPs of the viscosin group. Moreover, two of these cannot be discriminated from viscosin by MS/MS as they feature either a stereochemical difference (WLIP) or a Ile/Leu variation (massetolide F). Characterization of CLiPs should therefore typically be done by a complementary technique such as liquid state NMR spectroscopy. More so, it was recently described that the stereochemistry of a CLiP can be confirmed by NMR fingerprint matching (see below)


The structure of viscosin bears a single negative charge at physiological pH due to the presence of a glutamic acid at position 2. Consequently, the closest neutral isoform of viscosin is viscosinamide, which possesses a D-Gln2 instead of D-Glu2.
Three-dimensional structure
The three-dimensional structure of viscosin has not been described in literature. Although a viscosin crystal structure was originally published by Motley et al., this had to be corrected: the conformation possesses a D-Leu5, meaning that the compound is in fact the viscosin-isoform WLIP. (Motley, 2017)
Biophysical characteristics
Viscosin is a biosurfactant that can affect the surface tension of liquids in which it is dissolved. It can lower the water’s surface tension from 72 mN/m to 28 mN/m at a critical micellar concentration of 48 µM. (Saini, 2008) However, CMCs of 3.6-8.0 µM and 133 µM were also reported. (Hildebrand, 1998; Laycock, 1991)
Mode of action
Viscosin is able to permeabilise model membrane vesicles (PG:PE:cardiolipin and POPC). (Geudens, 2017) Similar to other viscosin group members, viscosin interacts both with the polar heads and aliphatic tails of model membrane systems, altering bilayer fluidity. The presence of cholesterol reduces the insertion depth of the viscosin molecules. Besides this, little to no data is available on the mode of action by which viscosin exerts its antagonistic activities against various microorganisms. However, there are a number of literature reports on pseudodesmin and WLIP; both members of the viscosin group, that likely possess a similar mode of action.
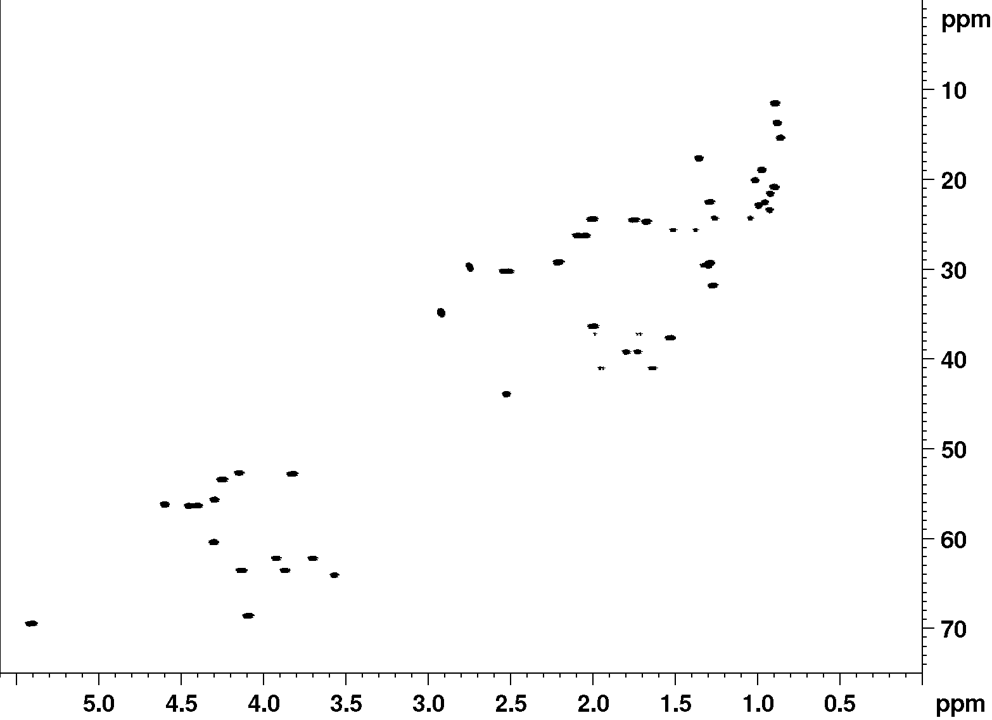
NMR fingerprint data
Recently, it was established that the planar structure and stereochemistry of CLiPs can be assessed by simple comparison to a reference. (De Roo, 2022) More specifically, by matching NMR spectra of a CLiP from a newly isolated bacterial source with those of existing (reference) CLiPs, one can determine whether they are identical or not. A detailed explanation on what NMR fingerprint matching is, and how to use it, can be found here.
Below, we provide the reference NMR data of viscosin in various formats. This data is recorded in DMF-d7 at room temperature, and can be used to asses similarities of newly isolated CLiPs to viscosin.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
- De Roo, et al. “An nuclear magnetic resonance fingerprint matching approach for the identification and structural re-evaluation of Pseudomonas lipopeptides.” Microbiology Spectrum 0, 0
https://dx.doi.org/doi:10.1128/spectrum.01261-22.
- Geudens, N., et al. (2014). ‘Impact of a stereocentre inversion in cyclic lipodepsipeptides from the viscosin group: a comparative study of the viscosinamide and pseudodesmin conformation and self-assembly’, ChemBioChem, 15: 2736-46.
https://doi.org/10.1002/cbic.201402389 - Geudens, N., et al. (2017). “Membrane interactions of natural cyclic lipodepsipeptides of the viscosin group.” Biochimica et Biophysica Acta – Biomembranes 1859(3): 331-339.
https://doi.org/10.1016/j.bbamem.2016.12.013 - Gerard, J., et al. (1997). “Massetolides A-H, antimycobacterial cyclic depsipeptides produced by two Pseumonads isolated from marine habitats.” Journal of Natural Products 60: 223-229.
https://doi.org/10.1021/np9606456 - Groupé, V., et al. (1951). “Observations on antiviral activity of viscosin.” Experimental Biology and Medicine 78(1): 354-358.
https://doi.org/10.3181/00379727-78-19071 - Hildebrand, P. D., et al. (1998). “Role of the biosurfactant viscosin in broccoli head rot caused by a pectolytic strain of Pseudomonas fluorescens.” Canadian Journal of Plant Pathology 20(3): 296-303.
https://doi.org/10.1021/jf00003a011 - Hiramoto, M., et al. (1969). “Synthesis of the proposed structure of viscosin.” Biochemical and Biophysical Research Communications 35(5): 702-706.
https://doi.org/10.1016/0006-291x(69)90462-8 - Hiramoto, M., et al. (1970). “The revised structure of viscosin, a peptide antibiotic.” Tetrahedron 13: 1087-1090.
https://doi.org/10.1016/S0040-4039(01)97915-8 - Laycock, M., et al. (1991). “Viscosin, a potent peptidolipid biosurfactant and phytopathogenic mediator produced by a pectolytic strain of Pseudomonas fluorescens.” Journal of Agricultural and Food Chemistry 39(3): 483-489.
https://doi.org/10.1021/jf00003a011 - Motley, et al. “Opportunistic sampling of roadkill as an entry point to accessing natural products assembled by bacteria associated with non-anthropoidal mammalian microbiomes.” Journal of Natural Products80, 3 (2017): https://dx.doi.org/10.1021/acs.jnatprod.6b00772.
- Saini, H. S., et al. (2008). “Efficient purification of the biosurfactant viscosin from Pseudomonas libanesis strain M9-3 and its physicochemical and biological properties.” Journal of Natural Products 71(6): 1011-1015.
https://doi.org/10.1021/np800069u