

Hoe kunnen we helpen?
WLIP
General info
| Original publication | Mortishire-Smith, 1991 |
| Original source | Pseudomonas fluorescens LMG 5329 (originally named Pseudomonas reactants) |
| Other known sources (non-putative) | Pseudomonas reactans NCPPB1311 (Lo Cantore, 2006) Pseudomonas putida RW10S2 (Rokni-Zadeh, 2012), Pseudomonas sp. (Motley, 2017) Pseudomonas sp. NSE1 (Omoboye, 2019), Pseudomonas sp. COW10 (Oni, 2020) |
| Stereochemistry determined by | X-ray crystallography: P. fluorescens LMG 5329 (Han, 1992), P. reactans NCPPB1311, Pseudomonas. sp. (Motley, 2017) NMR spectral matching: P. putida RW10S2, P. sp. NSE1, P. sp. COW10 |
Chemical properties
| CAS | 135096-89-8 |
| Molecular formula | C54H95N9O16 |
| Molecular weight | 1126.4 g/mol |
| Mono-isotopic mass | 1125.6897 Da (isobaric to other CLiPs, see text) |
| Solubility | Methanol, chloroform, acetone, buffer, DMF Not soluble (or very limited) in miliQwater |
| CMC | n.a. |
| Minimal surface tension | n.a. |
| 3D conformation | Crystal structure (Han, 1992, Motley, 2017) |
| NMR data available in literature | DMSO (Mortishire-Smith, 1991) DMF (Olorunleke, 2017) |
Introduction
WLIP (acronym for White Line Inducing Principle) was initially described as a secondary metabolite produced by a group of Pseudomonas species with the taxonomically incorrect designation Pseudomonas reactans. These bacteria are able to form a white line in agar when grown in the presence of the pathogen Pseudomonas tolaasii, the causal organism of brown blotch disease in mushrooms (Agaricus bisporus). The white line is a precipitate that is formed due to a specific interaction between diffusible compounds, more specifically WLIP (white line inducing principle/product) produced by Pseudomonas reactans and tolaasin, produced by Pseudomonas tolaasii. Both compounds are in fact cyclic lipodepsipeptides. Since then, it has been observed that the white-line-in-agar test is not specific, as other CLiPs can form a white line precipitate in the presence of tolaasin.
Biological activity
The antimicrobial activity of WLIP has often been analysed, with >160 assays available in literature. (reviewed in Geudens, 2018) Its activities are often described together with the antimicrobial activities of tolaasin, with which it forms a white line reaction. From these assays, it is clear that WLIP is mostly active against Gram-positive bacteria. In contrast, it is generally inactive against Gram-negative bacteria, with the notable exception of Xanthomonas species for which it does cause an antagonism. Against fungi, WLIP appears active in approx. 50% of the reported assays, although the strains are non-overlapping with those typically tested for other Pseudomonas CLiPs.
Chemical structure
The structure of WLIP was first elucidated in 1991 by means of mass spectrometry, liquid state NMR spectroscopy and chiral gas chromatography. (Mortishire-Smith, 1991). It consists of a 3-hydroxy decanoic acid (HDA) linked to a cyclic peptide consisting of 9 amino acids, of which 7 are contained in the macrocycle. More specifically, the chemical structure of WLIP is (R)-3-OH C10:0 – L-Leu1 – D-Glu2 – D-aThr3 – D-Val4 – D-Leu5 – D-Ser6 – L-Leu7 – D-Ser8 – L-Ile9. The molecule is cyclized by means of an ester bond between the C-terminus and the side chain hydroxyl moiety of threonine at position 3. The structure of WLIP bears a single negative charge at physiological pH due to the presence of a glutamic acid at position 2. The structure of WLIP belongs to the viscosin group, and strongly that of other group members such as pseudodesmin A, which differs only in the presence of Gln2 instead of Glu2, and viscosin, which differs only in the configuration of Leu5, being L-Leu5 in viscosin and D-Leu5 in WLIP.

The brute formula of WLIP – and therefore, its molecular mass – is identical to that of viscosin, massetolide F and massetolide L, all CLiPs of the viscosin group. Moreover, two of these cannot be discriminated from WLIP by MS/MS as they feature either a stereochemical difference (viscosin) or a Ile/Leu variation (massetolide F). Characterization of CLiPs should therefor typically be done by a complementary technique such as liquid state NMR spectroscopy.
Three dimensional conformation
The three-dimension structure of WLIP has independently been determined by means of liquid state NMR spectroscopy and X-ray crystallography. The solution structure, determined in dimethyl sulfoxide (DMSO), was described as resembling the seam of a tennis ball, whereby residues Ser6 to Ile9 appear to adopt a type II β-turn conformation. (Mortishire-Smith, 1991) Shortly after, the crystal structure was described. (Han, 1992) While the crystal structure did not show the presence of a β-turn between Ser6 to Ile9, it did adopt a single type II β-turn at the N-terminus, followed by a short left-handed helix ranging from D-Glu2 to D-Leu5. Comparison of the crystal structure with conformations of other viscosin group members shows high similarity (Sinnaeve, 2009), while the solution structure differs significantly. This is linked to the occurrence of conformational flexibility originating in the choice of DMSO as solvent. More specifically, DMSO has a very strong hydrogen bonding acceptor capability, out-competing the amide functionalities present in the peptidic part. Consequently, the solvent is a strong competitor for intramolecular hydrogen bond interactions and effectively acts as a denaturant for the cyclic peptide. The solution structure of WLIP should therefore be considered as irrelevant for any structure-activity relation.

Later, WLIP was also extracted from bacteria sampled from the ear canal of opossum roadkill. (Motley, 2017) Although first misidentified as viscosin, the conformation derived by x-ray crystallography revealed that it is in fact WLIP. Moreover, the crystal structure is identical to that reported earlier by Han et al.
NMR fingerprint data
Recently, it was established that the planar structure and stereochemistry of CLiPs can be assessed by simple comparison to a reference. (De Roo, 2022) More specifically, by matching NMR spectra of a CLiP from a newly isolated bacterial source with those of existing (reference) CLiPs, one can determine whether they are identical or not. A detailed explanation on what NMR fingerprint matching is, and how to use it, can be found here.
Below, we provide the reference NMR data of WLIP in various formats. This data is recorded in DMF-d7 at room temperature, and can be used to asses similarities of newly isolated CLiPs to WLIP.
{kind=link}
{kind=link}
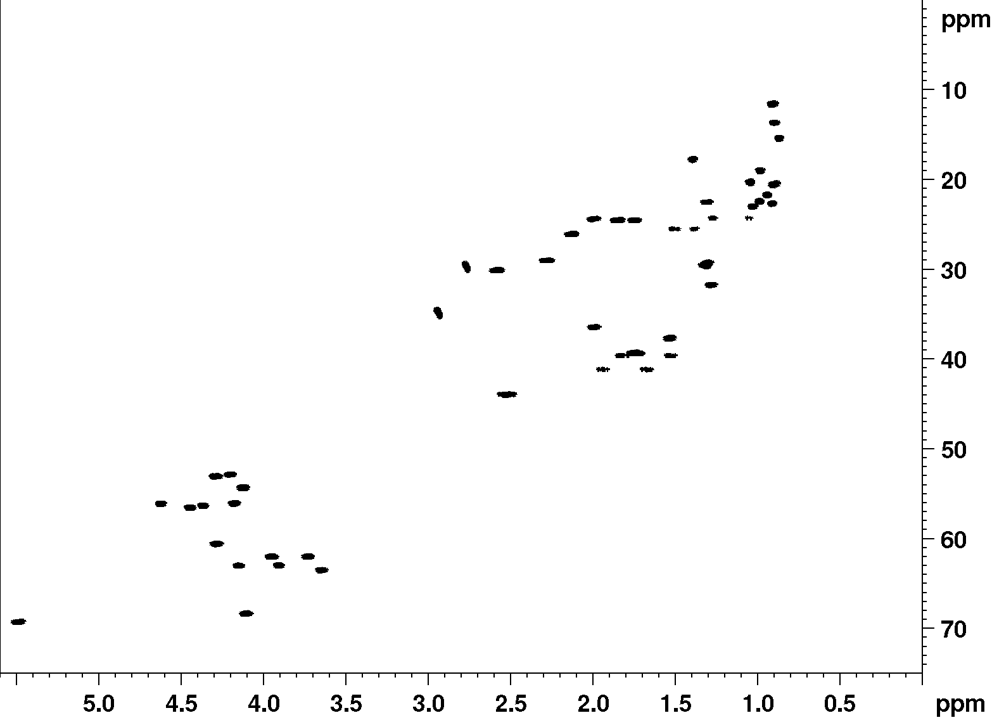{kind=link}
{kind=link}
References
De Roo, et al. “An nuclear magnetic resonance fingerprint matching approach for the identification and structural re-evaluation of Pseudomonas lipopeptides.” Microbiology Spectrum 0, 0 https://dx.doi.org/doi:10.1128/spectrum.01261-22
Geudens, et al. “Cyclic lipodepsipeptides from Pseudomonas spp. – Biological Swiss-Army Knives.” Frontiers in Microbiology9, 1867 (2018): https://dx.doi.org/10.3389/fmicb.2018.01867.
Han. “Structure of the White-Line-Inducing Principle isolated from Pseudomonas reactans.” Acta CrystallographicaC48 (1992).
Lo Cantore, et al. “Biological characterization of White Line-Inducing Principle (WLIP) produced by Pseudomonas reactans NCPPB1311.” Molecular Plant-Microbe Interactions19, 10 (2006): https://dx.doi.org/10.1094/mpmi.
Mortishire-Smith, et al. “Determination of the structure of an extracellular peptide produced by the mushroom Saprotroph Pseudomonas reactans.” Tetrahedron47, 22 (1991): https://dx.doi.org/10.1016/s0040-4020(01)80877-2.
Motley, et al. “Opportunistic sampling of roadkill as an entry point to accessing natural products assembled by bacteria associated with non-anthropoidal mammalian microbiomes.” Journal of Natural Products80, 3 (2017): https://dx.doi.org/10.1021/acs.jnatprod.6b00772.
Olorunleke. “Cyclic lipopeptides produced by Pseudomonas spp. associated with the cocoyam (Xanthosoma sagittifolium (L.) Schott) rhizosphere: diversity, regulation, secretion and biological activity.” Ghent University, 2017.
Omoboye, et al. “Pseudomonas sp. COW3 Produces New Bananamide-Type Cyclic Lipopeptides with Antimicrobial Activity against Pythium myriotylum and Pyricularia oryzae.” Molecules24, 22 (2019): https://dx.doi.org/10.3390/molecules24224170.
Oni, et al. “Cyclic lipopeptide-producing Pseudomonas koreensis group strains dominate the cocoyam rhizosphere of a Pythium root rot suppressive soil contrasting with P. putida prominence in conducive soils.” Environmental Microbiology22, 12 (2020): https://dx.doi.org/https://doi.org/10.1111/1462-2920.15127.
Rokni-Zadeh, et al. “Genetic and functional characterization of cyclic lipopeptide white-line-inducing principle (WLIP) production by rice rhizosphere isolate Pseudomonas putida RW10S2.” Applied and Environmental Microbiology78, 14 (2012): https://dx.doi.org/10.1128/AEM.00335-12.
Sinnaeve, et al. “The solution structure and self-association properties of the cyclic lipodepsipeptide pseudodesmin A support its pore-forming potential.” Chemistry – A European Journal15, 46 (2009): https://dx.doi.org/10.1002/chem.200901885.