

Hoe kunnen we helpen?
Orfamide A
Introduction
Orfamide A is a cyclic lipodepsipeptide produced by non-ribosomal peptide synthetases in various Pseudomonas species. It is the name-sake CLiP of a group of structurally similar lipopeptides, the orfamide group. This group also contains orfamides B, orfamide C (Gross, 2007), orfamides D – G (Ma, 2016), orfamide H (Ma, 2019), orfamides J – M ( De Roo, 2022), poaeamides A and B (Zachow, 2015, Nguyen, 2016) and the PPZPMs (Weisshoff, 2014).
The orfamides were first extracted from Pseudomonas protegens Pf-5 by employing a “genomisotopic approach” – a combination of genomic analysis and isotope-guided fractionation. Analysis of the orfamide structure was performed by an assay-guided approach to reveal the structure of orfamides A. (Gross, 2007) However, later it was found that the originally reported stereochemistry of orfamide A contains errors and should be revised to contain a D-Leu5 instead of L-Leu5 and a 3R-hydroxy fatty acid tail rather than a 3S-hydroxy one. (De Roo, 2022)
| Original publication | Gross, 2007 |
| Original source | Pseudomonas protegens Pf-5 |
| Sequence determined by | NMR spectroscopy, MS spectrometry |
| Stereochemistry determined by | Marfey’s analysis and chiral GC-MS (Gross, 2007) Corrected by NMR fingerprint matching and Marfey’s analysis (De Roo, 2022) |
| Other known sources (non-putative) | Pseudomonas protegens F6 (Jang, 2013), Pseudomonas sp. PH1b ( De Roo, 2022 ), Pseudomonas protegens CHA0 (De Roo, 2022) |
Chemical information
| CAS | 939960-34-6 |
| Molecular formula | C64H114N10O17 |
| Molecular weight | 1295.67 g/mol |
| Mono-isotopic mass | 1294.8363 Da |
| Solubility info | Soluble in methanol, ethanol, DMF, DMSO Limited solubility in acetonitrile |
| CMC | n.a. |
| 3D conformation | n.a. |
| NMR data available in literature | MeOH-d3 (Gross, 2007; De Roo, 2022) MeOH-d4 (Jang, 2013) DMF-d7 (Ma, 2016; De Roo, 2022) DMSO (Ma, 2019; De Roo, 2022) |
Characterisation
Orfamide A was initially characterized by detailed NMR analysis that allowed to determine the amino acid sequence and the presence of the N-terminal fatty acid moiety. The presence of the macrocycle between Thr3 CHβ and Val10 C’ was inferred from MS/MS analysis. Hydrolysis of orfamide A followed by chiral GC analysis and Marfey’s analysis confirmed the amino acids’ configuration. However, the originally published sequence contained errors in the stereochemical make-up of orfamide A. (Bando, 2022; De Roo, 2022) By means of NMR fingerprint matching and Marfey’s analysis, it was shown that the sequence contains a D-Leu5 and an 3R-hydroxyfatty acid. (De Roo, 2022) Independently, Bando et al. can to the same conclusion using a solid-state peptide synthesis approach. (Bando, 2022)) Consequently, the corrected structure of orfamide A is 3R-OH C14:0 – L-Leu1 – D-Glu2 – D-aThr3 – D-aIle4 – D-Leu5 – D-Ser6 – L-Leu7 – L-Leu8 – D-Ser9 – L-Val10 and is cyclized by means of an ester bond between the C-terminal carbonyl and the hydroxyl side chain of D-alloThr3. Orfamide A is the name-sake of the orfamide group, a collection of structurally similar lipopeptides.


Since it first description, several other non-putative orfamide A producers are described. For example, orfamide A was isolated from P. protegens F6 during a screening assay of biosurfactant-producing bacteria. (Jang, 2013) By matching the NMR spectra of F6 with those of orfamide A produced by the original producer P. protegens Pf-5, it was concluded that both compounds are identical. (Jang, 2013; De Roo, 2022)
Based on the similarity of the NRPS with that of Pseudomonas protegens Pf-5, it was hypothesized that Pseudomonas sp. PH1b produces orfamide A. (Ma, 2016) Recently, this was independently confirmed by means of a novel NMR fingerprint matching procedure. (De Roo, 2022)
Pseudomonas sp. CHA0 was reported to produce an structural analogue of orfamide A, named orfamide H, differing in the length of the fatty acid tail. (Ma, 2019) Using NMR spectroscopy, it was established that orfamide H possesses an identical amino acid sequence, but would differ in the presence of a non-branched 3-hydroxyl pentadecanoic fatty acid (3-OH C15:0). However, an error occurred in the NMR assignment, whereby the additional methylene (CH2) unit is not associated with the fatty acid residue, but originates from a Fisher esterification reaction of D-Glu2 with methanol (used as solvent during the extraction procedure) resulting in a COOCH3 side chain moiety rather than COOH. Since this is the result of the extraction procedure, rather than post-modification by the producing bacterium, P. protegens CHA0 does in fact produce orfamide A.
Bioactivity
Orfamide A showed antagonistic activity against plant pathogens such as Phytophthora strains (porri and ramorum), Pythium ultimum and Rhizoctonia solani. (Gross, 2007, Ma, 2016) In contrast, orfamide H, the methylester of orfamide A, did not show any antibiotic activities against human pathogens such as Staphylococcus aureus, Escherichia coli, and Salmonella typhimurium, or plant pathogens like Magnaporthe oryzae, Botrytis cinerea, Rhizoctonia solani, and Fusarium graminearum. (Ma, 2019) Therefor, it is likely that the introduction of a methyl ester on the side chain of D-Glu2 disrupts the antimicrobial activities of orfamide A.
Orfamide A has not been tested against Gram-positive or Gram-negative bacteria. (Geudens, 2018)
The orfamides are among the few CLiPs that been investigated for their insecticidal activities: it is found that orfamide A shows a dose-dependent insecticidal activity against green peach aphid (Myzus persicae). (Jang, 2013)
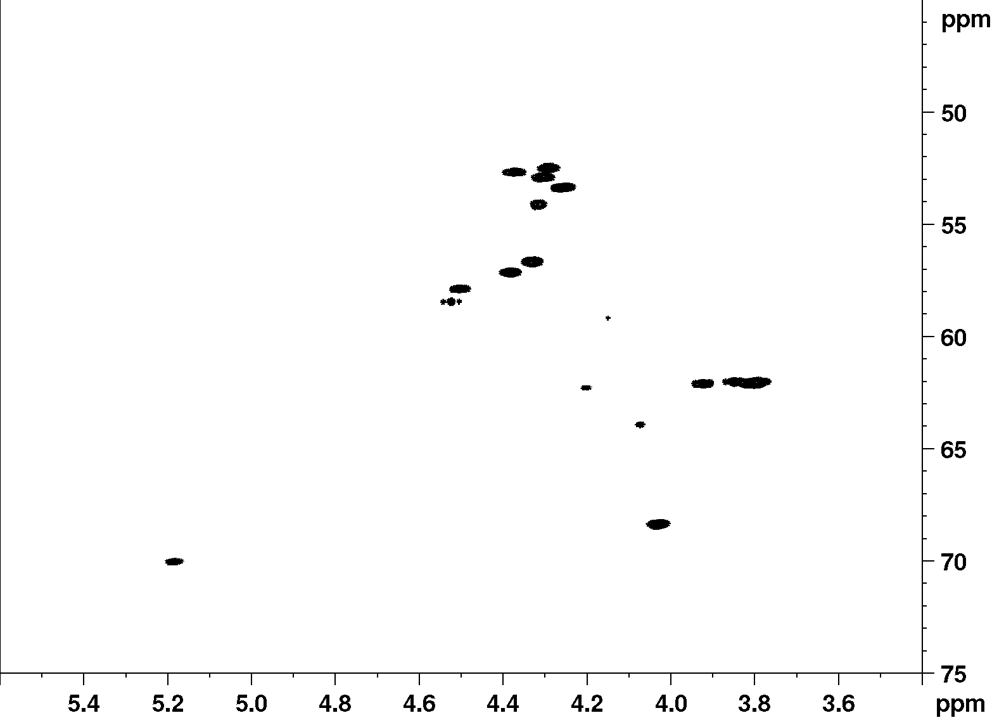
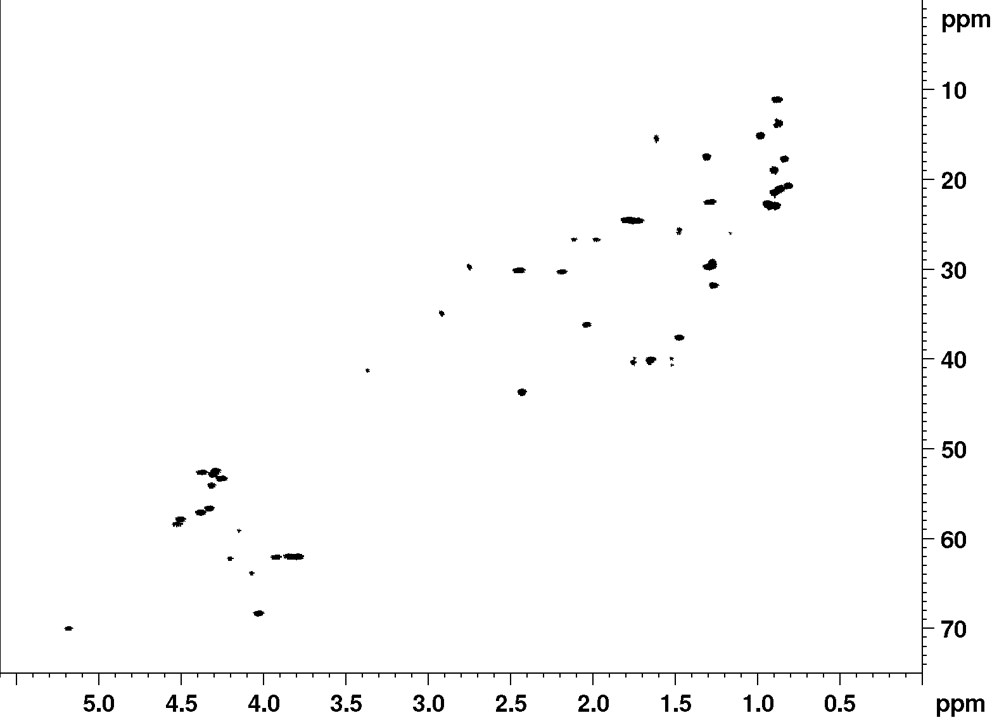
NMR fingerprint data
Recently, it was established that the planar structure and stereochemistry of CLiPs can be assessed by simple comparison to a reference. (De Roo, 2022) More specifically, by matching NMR spectra of a CLiP from a newly isolated bacterial source with those of existing (reference) CLiPs, one can determine whether they are identical or not. A detailed explanation on what NMR fingerprint matching is, and how to use it, can be found here.
Below, we provide the reference NMR data of orfamide A in various formats. This data is recorded in DMF-d7 at room temperature, and can be used to asses similarities of newly isolated CLiPs to orfamide A.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
Bando, et al. “Total Synthesis and Structure Correction of the Cyclic Lipodepsipeptide Orfamide A.” Chemistry – A European Journal 28, 20 (2022):
https://dx.doi.org/https://doi.org/10.1002/chem.202104417.
De Roo, et al. “An nuclear magnetic resonance fingerprint matching approach for the identification and structural re-evaluation of Pseudomonas lipopeptides.” Microbiology Spectrum 0, 0
https://dx.doi.org/doi:10.1128/spectrum.01261-22.
Geudens, et al. “Cyclic lipodepsipeptides from Pseudomonas spp. – Biological Swiss-Army Knives.” Frontiers in Microbiology9, 1867 (2018): https://dx.doi.org/10.3389/fmicb.2018.01867.
Gross, et al. “The genomisotopic approach: a systematic method to isolate products of orphan biosynthetic gene clusters.” Chemistry & Biology14, 1 (2007): https://dx.doi.org/10.1016/j.chembiol.2006.11.007.
Jang, et al. “Identification of orfamide A as an insecticidal metabolite produced by Pseudomonas protegens F6.” Journal of Agricultural and Food Chemistry61, 28 (2013): https://dx.doi.org/10.1021/jf401218w.
Ma, et al. “Biosynthesis, chemical structure, and structure-activity relationship of orfamide lipopeptides produced by Pseudomonas protegens and related species.” Frontiers in Microbiology7 (2016): https://dx.doi.org/10.3389/fmicb.2016.00382.
Ma, et al. “Isolation and characterization of a new cyclic lipopeptide orfamide H from Pseudomonas protegens CHA0.” Journal of Antibiotics (2019): https://dx.doi.org/10.1038/s41429-019-0254-0.
Nguyen, et al. “Indexing the Pseudomonas specialized metabolome enabled the discovery of poaeamide B and the bananamides.” Nature Microbiology2 (2016): https://dx.doi.org/10.1038/nmicrobiol.2016.197.
Weisshoff, et al. “PPZPMs – a novel group of cyclic lipodepsipeptides produced by the Phytophtora alni associated strain Pseudomonas sp. JX090307 – the missing link between the viscosin and amphisin group.” Natural Products Communications9, 7 (2014).
Zachow, et al. “The novel lipopeptide poaeamide of the endophyte Pseudomonas poae RE*1-1-14 is involved in pathogen suppression and root colonization.” Molecular Plant-Microbe Interactions28, 7 (2015): https://dx.doi.org/10.1094/MPMI-12-14-0406-R.